14.8 The Endomembrane System
Elizabeth Dahlhoff
Learning Objectives
By the end of this section, you will be able to do the following:
- Explain the role of the endomembrane system in protein modification, packaging, and intracellular transport.
- Describe the relationship between the endomembrane system structure and its functions.
After Translation: Protein Trafficking Through the Endomembrane System
Although the translation of mRNAs whose protein products function in the cytoplasm takes place on free-floating ribosomes, there are a subset of mRNAs that must go through a slightly modified translation process, namely those that code for membrane or secreted proteins or proteins that function in the lumen of organelles that make up the endomembrane system. Through their interactions with the endomembrane system, these proteins are trafficked (translocated) to their ultimate destination in (or out of) the cell.
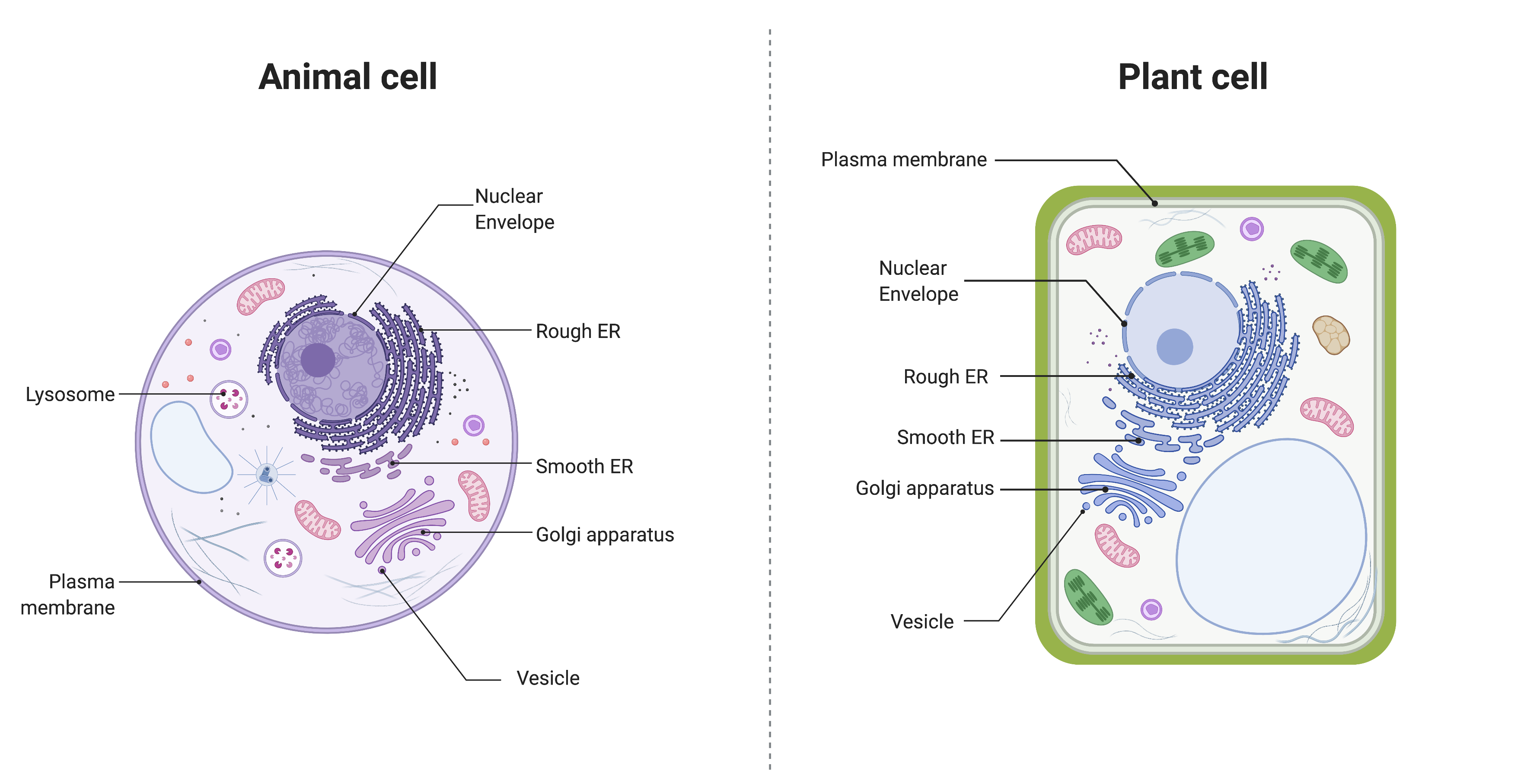
The endomembrane system (endo = “within”) is a group of membranes and organelles in eukaryotic cells that works together to modify, package, and transport lipids and proteins. It includes the nuclear envelope, lysosomes, vesicles, endoplasmic reticulum, and Golgi apparatus. Although not technically within the cell, the plasma membrane is included in the endomembrane system because it interacts with the other endomembranous organelles. You can review the structure and key features of the plasma membrane here. The endomembrane system does not include either mitochondria or chloroplast membranes.
The Endoplasmic Reticulum
The endoplasmic reticulum (ER) is a series of interconnected membranous sacs and tubules that collectively modifies proteins and synthesizes lipids. However, these two functions take place in separate areas of the ER: the rough ER and the smooth ER, respectively.
We call the interior portion of the ER the lumen. The ER’s membrane, which is a phospholipid bilayer embedded with proteins, is continuous with the nuclear envelope.

Rough ER
Scientists have named the rough endoplasmic reticulum (RER) as such because the ribosomes attached to its cytoplasmic surface give it a studded appearance when viewing it through an electron microscope.
Ribosomes can be classified as either free or membrane-bound. Free ribosomes exist in the cytosol and are not associated with a membrane. Membrane-bound ribosomes are attached to the outside of the membrane of the endoplasmic reticulum. Free and membrane-bound ribosomes are identical in structure; they differ only in location. In fact, an individual ribosome may sometimes be free and sometimes be membrane-bound, depending on what type of protein it is producing.
Free ribosomes generally produce proteins that will be used within the cytoplasm. Membrane-bound ribosomes generally produce proteins that will either be embedded in a membrane, or secreted from the cell.
All ribosomes begin translation as free ribosomes. However, if they produce a stretch of hydrophobic amino acids, they are triggered to dock onto specialized pores on the RER membrane and transfer their newly synthesized proteins into the RER’s lumen where they undergo structural modifications, such as folding or acquiring side chains. These modified proteins incorporate into cellular membranes—such as the ER membrane, the membranes of other organelles, or the cell membrane. The proteins can also be secreted from the cell (such as protein hormones, enzymes). The RER also makes phospholipids for cellular membranes.
If the phospholipids or modified proteins are not destined to stay in the RER, they will reach their destinations via transport vesicles that bud from the RER’s membrane.
Since the RER is engaged in modifying proteins (such as enzymes, for example) that secrete from the cell, you would be correct in assuming that the RER is abundant in cells that secrete proteins. This is the case with liver cells, for example.
Smooth ER
The smooth endoplasmic reticulum (SER) is continuous with the RER but has few or no ribosomes on its cytoplasmic surface. SER functions include synthesis of carbohydrates, lipids, and steroid hormones; detoxification of medications and poisons; and storing calcium ions. In muscle cells, a specialized SER, the sarcoplasmic reticulum, is responsible for storing calcium ions that are needed to trigger the muscle cells’ coordinated contractions.
The Golgi Apparatus
We have already mentioned that vesicles can bud from the ER and transport their contents elsewhere, but where do the vesicles go? Before reaching their final destination, the lipids or proteins within the transport vesicles still need sorting, packaging, and tagging so that they end up in the right place. Sorting, tagging, packaging, and distributing lipids and proteins takes place in the Golgi apparatus (also called the Golgi body), a series of flattened membranous sacs.
The side of the Golgi apparatus that is closer to the ER is called the cis face. The opposite side is the trans face. The transport vesicles that formed from the ER travel to the cis face, fuse with it, and empty their contents into the Golgi apparatus’ lumen. As the proteins and lipids travel through the Golgi, they undergo further modifications that allow them to be sorted. The most frequent modification is adding short sugar molecule chains. These newly modified proteins and lipids are then tagged with phosphate groups or other small molecules in order to travel to their proper destinations.
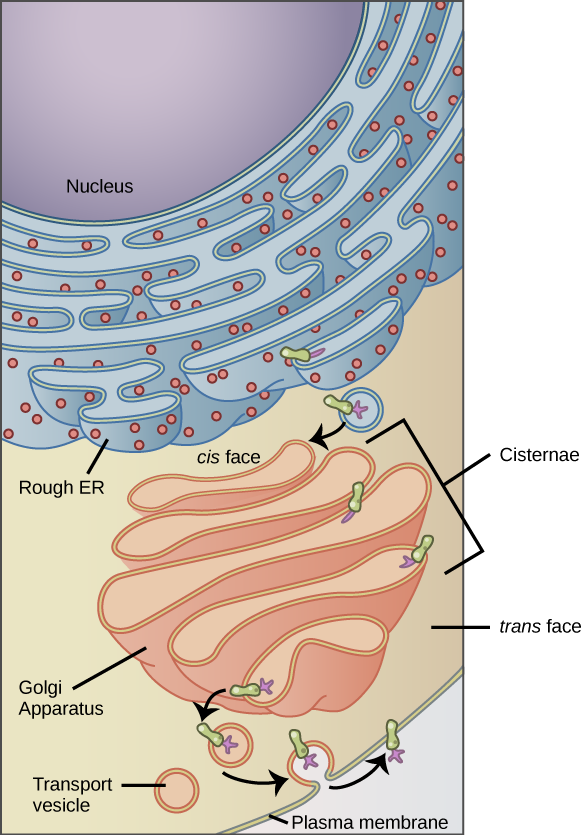
Finally, the modified and tagged proteins are packaged into transport vesicles that bud from the Golgi’s trans face. While some of these vesicles deposit their contents into other cell parts where they will be used, other secretory vesicles fuse with the plasma membrane and release their contents outside the cell.
In another example of form following function, cells that engage in a great deal of secretory activity (such as salivary gland cells that secrete digestive enzymes or immune system cells that secrete antibodies) have an abundance of Golgi. In plant cells, the Golgi apparatus has the additional role of synthesizing polysaccharides, some of which are incorporated into the cell wall and some of which other cell parts use.
Lysosomes
Lysosomes are part of the endomembrane system. Lysosomes are found in animal cells. They contain hydrolytic enzymes that are used to digest macromolecules and worn-out organelles.
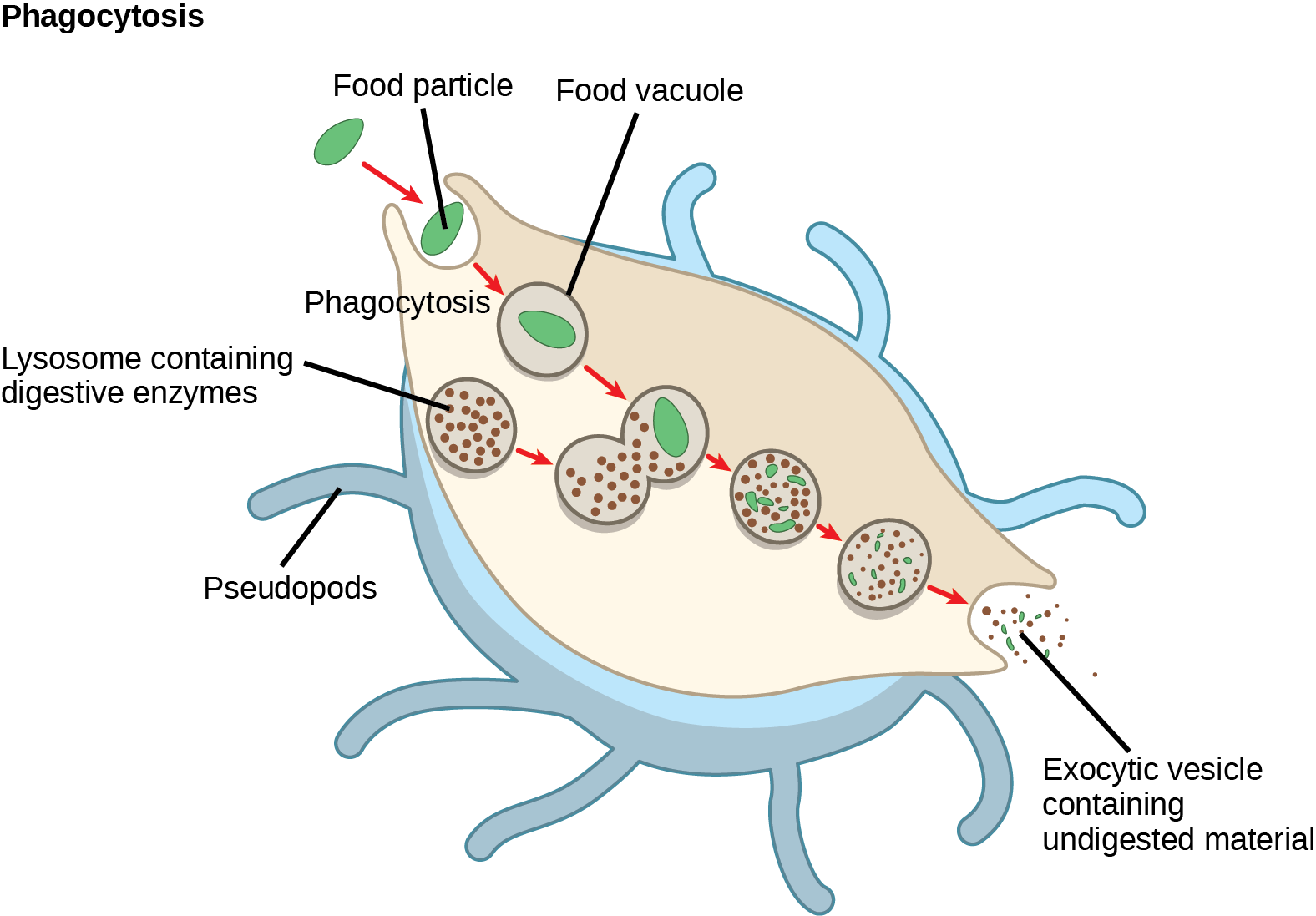
Lysosomes also use their hydrolytic enzymes to destroy pathogens (disease-causing organisms) that might enter the cell. A good example of this occurs in macrophages, a group of white blood cells which are part of your body’s immune system. In a process that scientists call phagocytosis or endocytosis, a section of the macrophage’s plasma membrane invaginates (folds in) and engulfs a pathogen. The invaginated section, with the pathogen inside, then pinches itself off from the plasma membrane and becomes a vesicle. The vesicle fuses with a lysosome. The lysosome’s hydrolytic enzymes then destroy the pathogen.
Trafficking
Protein trafficking is the process by which proteins are directed to specific locations within or outside the cell after they are synthesized. This process begins in the ribosomes, where proteins are made. Proteins destined for secretion or for use in organelles like the lysosome or plasma membrane are usually synthesized by ribosomes on the rough endoplasmic reticulum (RER). As they are made, these proteins enter the RER, where they undergo folding and modification. Next, the proteins are transported in vesicles to the Golgi apparatus, where they are further modified, sorted, and packaged. Based on molecular signals, the Golgi directs each protein to its correct destination—this could be the plasma membrane, lysosomes, other organelles, or outside the cell via exocytosis. Proteins that are made by free ribosomes in the cytosol generally stay in the cytoplasm or go to the nucleus, mitochondria, or peroxisomes, guided by signal sequences within the protein itself.
Here is a video summarizing the process.
Video 14.8.1. Protein Trafficking by ndsuvirtualcell
Traffic(king) check
Section Summary
The endomembrane system includes the nuclear envelope, lysosomes, vesicles, the ER, and Golgi apparatus, as well as the plasma membrane. These cellular components work together to modify, package, tag, and transport proteins and lipids that form the membranes. The RER modifies proteins and synthesizes phospholipids used in cell membranes. The SER synthesizes carbohydrates, lipids, and steroid hormones; engages in the detoxification of medications and poisons; and stores calcium ions. Sorting, tagging, packaging, and distribution of lipids and proteins take place in the Golgi apparatus. Lysosomes are created by the budding of the membranes of the RER and Golgi. Lysosomes digest macromolecules, recycle worn-out organelles, and destroy pathogens. These organelles all play a key role in protein trafficking.
Practice Questions
Glossary
- endomembrane system
- group of organelles and membranes in eukaryotic cells that work together modifying, packaging, and transporting lipids and proteins
- endoplasmic reticulum (ER)
- series of interconnected membranous structures within eukaryotic cells that collectively modify proteins and synthesize lipids
- Golgi apparatus
- eukaryotic organelle made up of a series of stacked membranes that sorts, tags, and packages lipids and proteins for distribution
- rough endoplasmic reticulum (RER)
- region of the endoplasmic reticulum that is studded with ribosomes and engages in protein modification and phospholipid synthesis
- smooth endoplasmic reticulum (SER)
- region of the endoplasmic reticulum that has few or no ribosomes on its cytoplasmic surface and synthesizes carbohydrates, lipids, and steroid hormones; detoxifies certain chemicals (like pesticides, preservatives, medications, and environmental pollutants), and stores calcium ions
Figure Descriptions
Figure 14.8.1. The image compares the endomembrane system in an animal cell (left) and a plant cell (right). Both diagrams label the nuclear envelope, rough endoplasmic reticulum (ER), smooth ER, Golgi apparatus, vesicles, and plasma membrane. The animal cell is shown as a round cell with additional labeled lysosomes, while the plant cell is rectangular with a thick green cell wall and contains large central vacuole, chloroplasts, and other organelles in addition to the labeled endomembrane components. [Return to Figure 14.8.1]
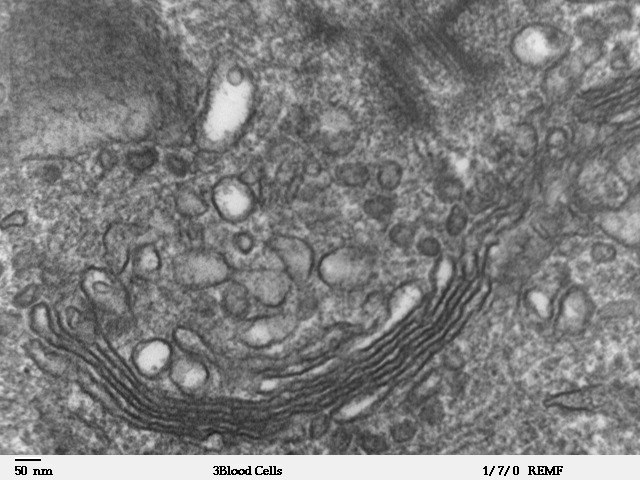
Figure 14.8.2. The composite image presents the endoplasmic reticulum in different views. Part (a) is a color illustration showing the cell nucleus on the left with rough ER extending outward, studded with small red ribosomes, and transitioning into smooth ER represented as smooth tubular membranes without ribosomes. Part (b) is a high-magnification electron micrograph (×110,000) showing stacked layers of rough ER with ribosomes attached, appearing as small dark dots. Part (c) is another high-magnification electron micrograph (×110,510) showing smooth ER as a network of tubular membranes without ribosomes, located beneath the cell membrane and adjacent to the nucleus. [Return to Figure 14.8.2]
Figure 14.8.4. A diagram showing the pathway of membrane and secretory proteins from the rough endoplasmic reticulum (RER) to the Golgi apparatus and then to the plasma membrane. The RER, located next to the nucleus, is shown with ribosomes on its surface. A green integral membrane protein, modified by a purple carbohydrate, buds off from the RER in a vesicle and fuses with the cis face of the Golgi apparatus. Inside the Golgi’s stacked cisternae, the protein is further modified with additional carbohydrates. A vesicle containing the completed protein buds from the trans face of the Golgi and moves toward the plasma membrane. When the vesicle fuses with the membrane, the protein becomes embedded in the membrane. [Return to Figure 14.8.4]
Figure 14.8.5. A macrophage is illustrated engulfing a green bacterium through a process labeled “cytosis.” The bacterium is enclosed in a vesicle, which fuses with vesicles containing brown lysosomes. The bacterium is gradually broken down into smaller pieces inside the vesicles. The digested material is then released from the macrophage. Other organelles are not shown. [Return to Figure 14.8.5]
Licenses and Attributions
Media Attributions
- Endomembrane System © Melissa Hardy is licensed under a CC BY-NC (Attribution NonCommercial) license
- Human leukocyte showing golgi © Louisa Howard is licensed under a Public Domain license
- Golgi protein © OpenStax is licensed under a CC BY (Attribution) license
- lysosome © OpenStax is licensed under a CC BY (Attribution) license

{kind=link}