Chapter 3. The Cellular Level of Organization
3.1 The Cytoplasm and Cellular Organelles
Learning Objectives
By the end of this section, you will be able to:
- Describe the structure and function of the cellular organelles associated with the endomembrane system, including the endoplasmic reticulum, Golgi apparatus, and lysosomes
- Describe the structure and function of mitochondria and peroxisomes
- Explain the three components of the cytoskeleton, including their composition and functions
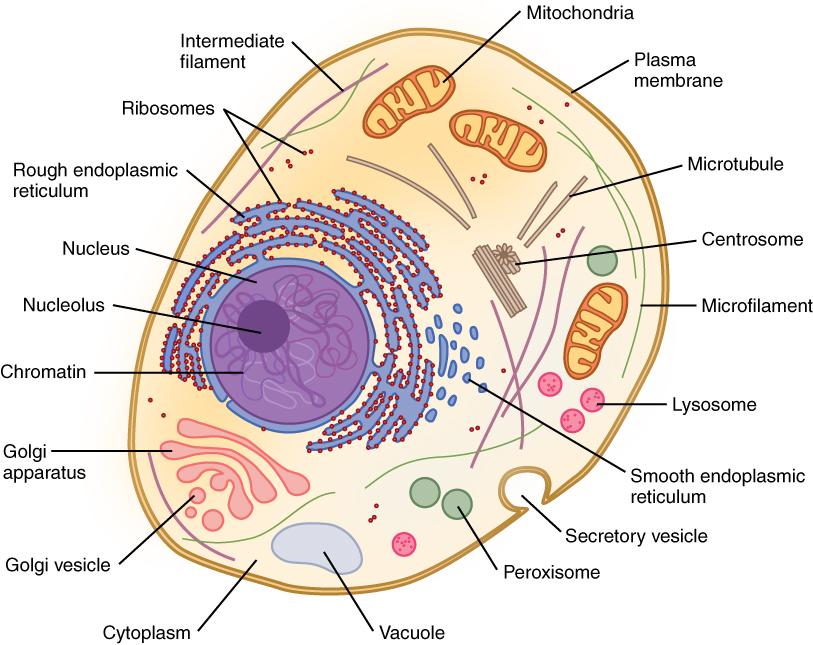
Now that you have learned that the cell membrane surrounds all cells, you can dive inside of a prototypical human cell to learn about its internal components and their functions. All living cells in multicellular organisms contain an internal cytoplasmic compartment and a nucleus within the cytoplasm. Cytosol, the jelly-like substance within the cell, provides the fluid medium necessary for biochemical reactions. Eukaryotic cells, including all animal cells, also contain various cellular organelles. An organelle (“little organ”) is one of several different types of membrane-enclosed bodies in the cell, each performing a unique function. Just as the various bodily organs work together in harmony to perform all of a human’s functions, the many different cellular organelles work together to keep the cell healthy and performing all of its important functions. The organelles and cytosol, taken together, compose the cell’s cytoplasm. The nucleus is a cell’s central organelle, which contains the cell’s DNA (Figure 3.1.1).
Organelles of the Endomembrane System
A set of three major organelles together form a system within the cell called the endomembrane system. These organelles work together to perform various cellular jobs, including the task of producing, packaging, and exporting certain cellular products. The organelles of the endomembrane system include the endoplasmic reticulum, Golgi apparatus, and vesicles.
Endoplasmic Reticulum
The endoplasmic reticulum (ER) is a system of channels that is continuous with the nuclear membrane (or “envelope”) covering the nucleus and composed of the same lipid bilayer material. The ER can be thought of as a series of winding thoroughfares similar to the waterway canals in Venice. The ER provides passages throughout much of the cell that function in transporting, synthesizing, and storing materials. The winding structure of the ER results in a large membranous surface area that supports its many functions (Figure 3.1.2).
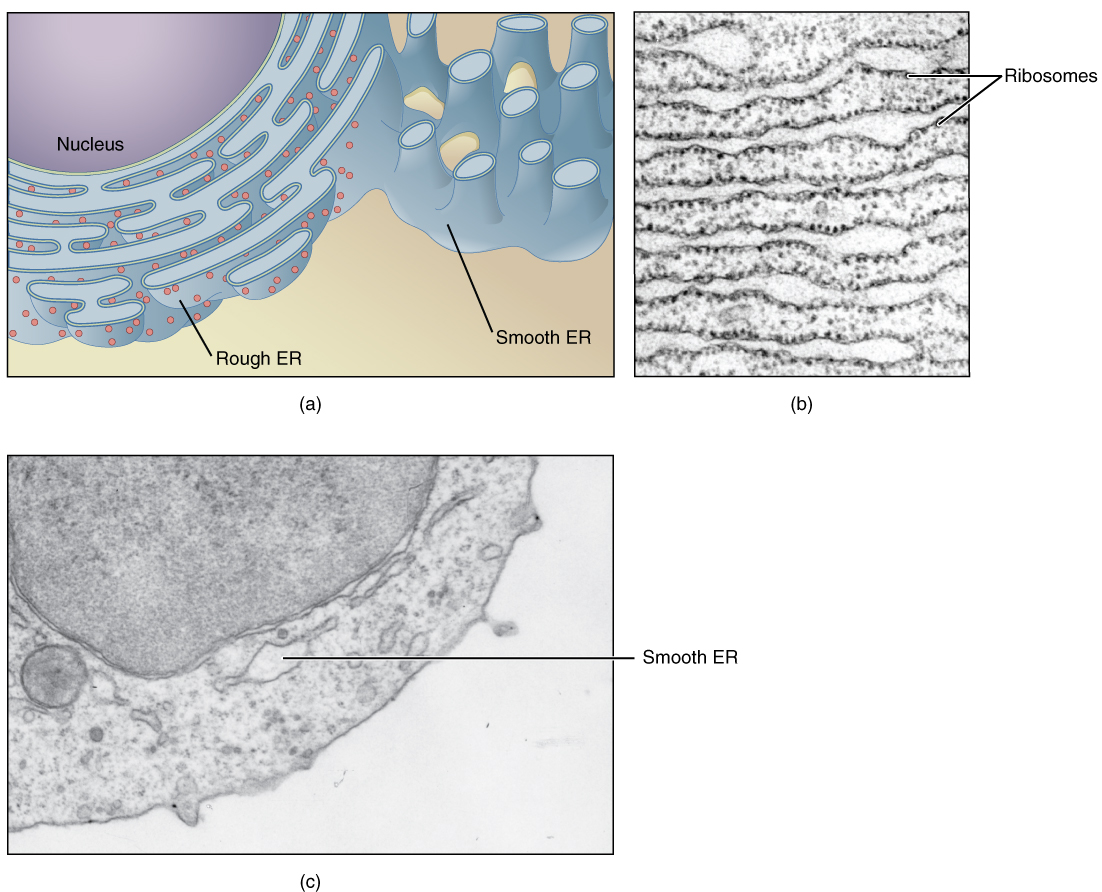
Endoplasmic reticulum can exist in two forms: rough ER and smooth ER. These two types of ER perform some very different functions and can be found in very different amounts depending on the type of cell. Rough ER is so-called because its membrane is dotted with embedded granules—organelles called ribosomes, giving the rough ER a bumpy appearance. A ribosome is an organelle that serves as the site of protein synthesis. It is composed of two ribosomal RNA subunits that wrap around mRNA to start the process of translation, followed by protein synthesis. Smooth ER lacks these ribosomes.
One of the main functions of the smooth ER is in the synthesis of lipids. The smooth ER synthesizes phospholipids, the main component of biological membranes, as well as steroid hormones. For this reason, cells that produce large quantities of such hormones, such as those of the female ovaries and male testes, contain large amounts of smooth ER. In addition to lipid synthesis, the smooth ER also sequesters (i.e., stores) and regulates the concentration of cellular Ca++, a function extremely important in cells of the nervous system where Ca++ is the trigger for neurotransmitter release. The smooth ER additionally metabolizes some carbohydrates and performs a detoxification role, breaking down certain toxins.
In contrast with the smooth ER, the primary job of the rough ER is the synthesis and modification of proteins destined for the cell membrane or for export from the cell. For this protein synthesis, many ribosomes attach to the ER (giving it the studded appearance of rough ER). Typically, a protein is synthesized within the ribosome and released inside the channel of the rough ER, where sugars can be added to it (by a process called glycosylation) before it is transported within a vesicle to the next stage in the packaging and shipping process: the Golgi apparatus.
The Golgi Apparatus
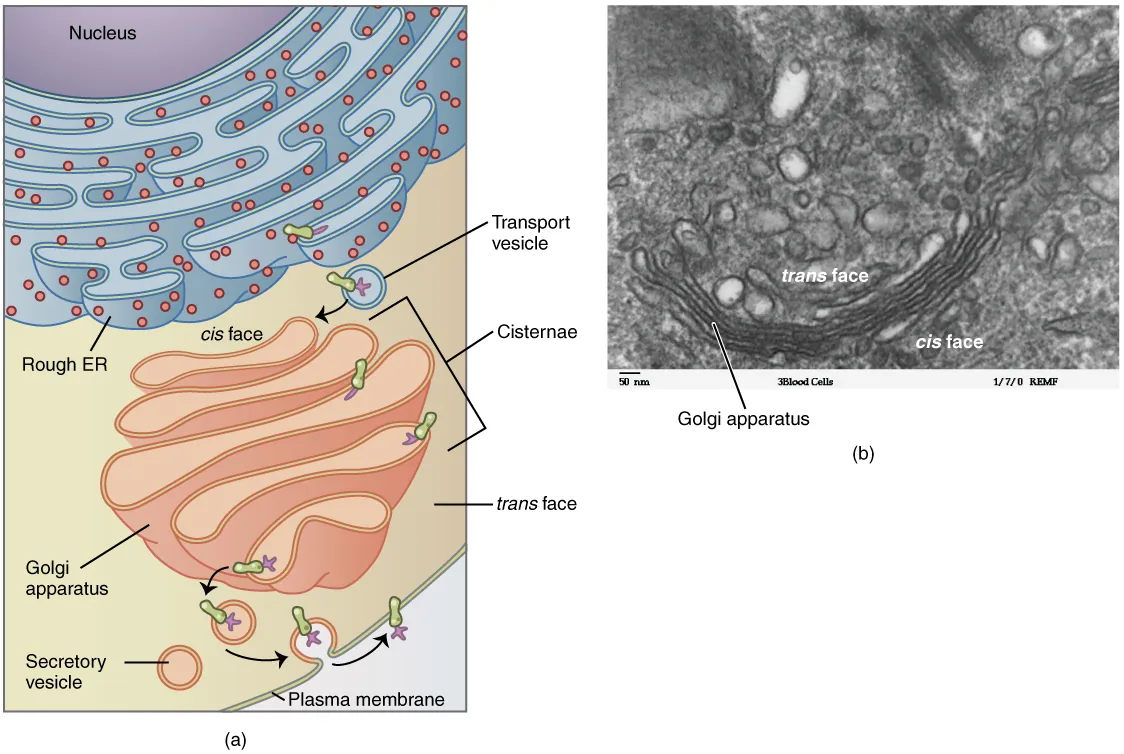
The Golgi apparatus is responsible for sorting, modifying, and shipping off the products that come from the rough ER, much like a post office. The Golgi apparatus looks like stacked flattened discs, almost like stacks of oddly shaped pancakes. Like the ER, these discs are membranous. The Golgi apparatus has two distinct sides, each with a different role. One side of the apparatus receives products in vesicles. These products are sorted through the apparatus and then they are released from the opposite side after being repackaged into new vesicles. If the product is to be exported from the cell, the vesicle migrates to the cell surface and fuses to the cell membrane, and the cargo is secreted (Figure 3.1.3).
Lysosomes
Some of the protein products packaged by the Golgi include digestive enzymes that are meant to remain inside the cell for use in breaking down certain materials. The enzyme-containing vesicles released by the Golgi may form new lysosomes, or fuse with existing lysosomes. A lysosome is an organelle that contains enzymes that break down and digest unneeded cellular components, such as a damaged organelle. (A lysosome is similar to a wrecking crew that takes down old and unsound buildings in a neighborhood.) Autophagy (“self-eating”) is the process of a cell digesting its own structures. Lysosomes are also important for breaking down foreign material. For example, when certain immune defense cells (white blood cells) phagocytize bacteria, the bacterial cell is transported into a lysosome and digested by the enzymes inside. As one might imagine, such phagocytic defense cells contain large numbers of lysosomes.
Under certain circumstances, lysosomes perform a more grand and dire function. In the case of damaged or unhealthy cells, lysosomes can be triggered to open up and release their digestive enzymes into the cytoplasm of the cell, killing the cell. This “self-destruct” mechanism is called autolysis, and makes the process of cell death controlled (a mechanism called “apoptosis”).
Organelles for Energy Production and Detoxification
In addition to the jobs performed by the endomembrane system, the cell has many other important functions. Just as you must consume nutrients to provide yourself with energy, so must each of your cells take in nutrients, some of which convert to chemical energy that can be used to power biochemical reactions. Another important function of the cell is detoxification. Humans take in all sorts of toxins from the environment and also produce harmful chemicals as byproducts of cellular processes. Cells called hepatocytes in the liver detoxify many of these toxins.
Mitochondria
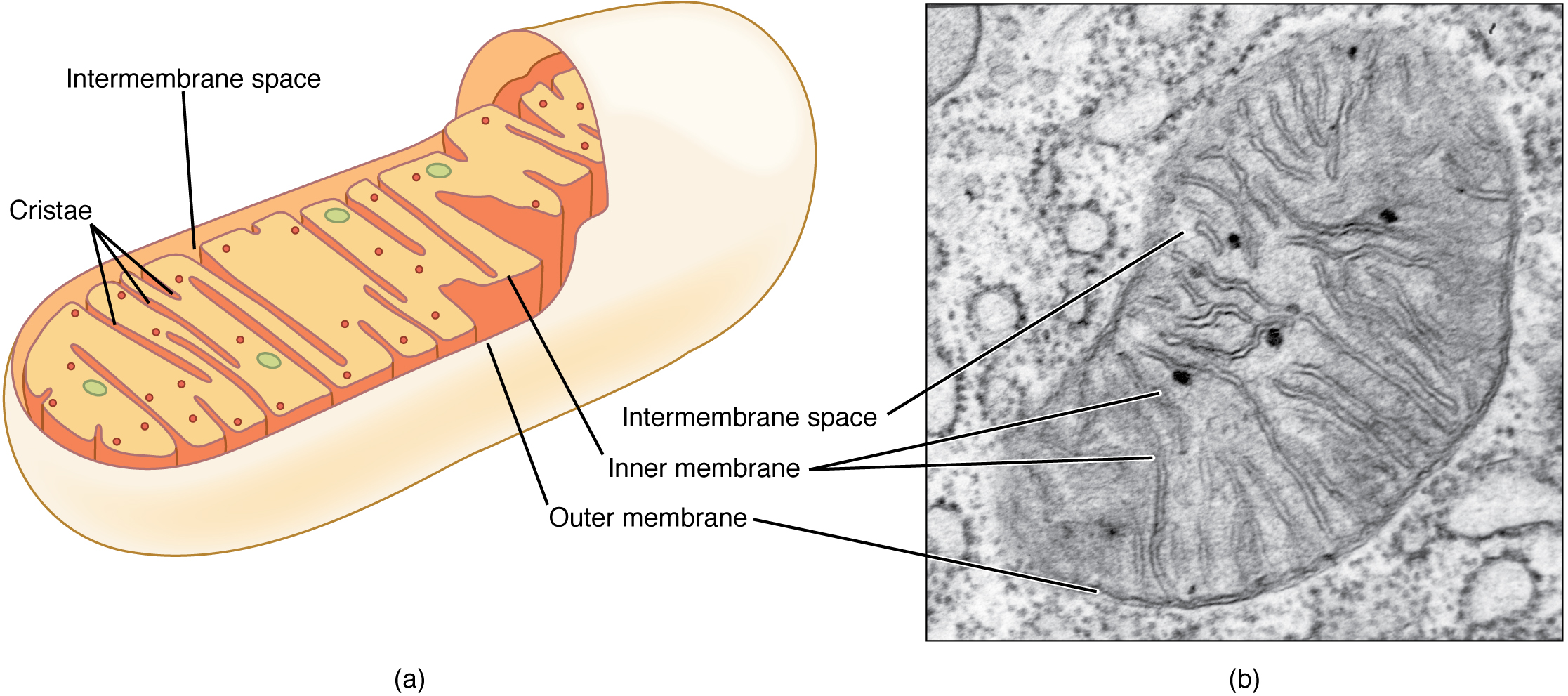
A mitochondrion (plural mitochondria) is a membranous, bean-shaped organelle that is the “energy transformer” of the cell. Mitochondria consist of an outer lipid bilayer membrane as well as an additional inner lipid bilayer membrane (Figure 3.1.4). The inner membrane is highly folded into winding structures with a great deal of surface area, called cristae. It is along this inner membrane that a series of proteins, enzymes, and other molecules perform the biochemical reactions of cellular respiration. These reactions convert energy stored in nutrient molecules (such as glucose) into adenosine triphosphate (ATP), which provides usable cellular energy to the cell. Cells use ATP constantly, so the mitochondria are constantly at work. Oxygen molecules are required during cellular respiration, which is why you must constantly breathe it in. One of the organ systems in the body that uses huge amounts of ATP is the muscular system because ATP is required to sustain muscle contraction. As a result, muscle cells are packed full of mitochondria. Nerve cells also need large quantities of ATP to run their sodium-potassium pumps. Therefore, an individual neuron will be loaded with over a thousand mitochondria. On the other hand, a bone cell, which is not nearly as metabolically active, might only have a couple hundred mitochondria.
Peroxisomes
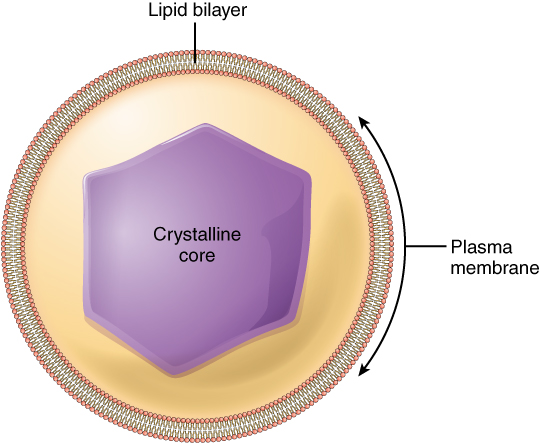
Like lysosomes, a peroxisome is a membrane-bound cellular organelle that contains mostly enzymes (Figure 3.1.5). Peroxisomes perform a couple of different functions, including lipid metabolism and chemical detoxification. In contrast to the digestive enzymes found in lysosomes, the enzymes within peroxisomes serve to transfer hydrogen atoms from various molecules to oxygen, producing hydrogen peroxide (H2O2). In this way, peroxisomes neutralize poisons such as alcohol. In order to appreciate the importance of peroxisomes, it is necessary to understand the concept of reactive oxygen species.
Reactive oxygen species (ROS) such as peroxides and free radicals are the highly reactive products of many normal cellular processes, including the mitochondrial reactions that produce ATP and oxygen metabolism. Examples of ROS include the hydroxyl radical OH, H2O2, and superoxide (O2−). Some ROS are important for certain cellular functions, such as cell signaling processes and immune responses against foreign substances. Free radicals are reactive because they contain free unpaired electrons; they can easily oxidize other molecules throughout the cell, causing cellular damage and even cell death. Free radicals are thought to play a role in many destructive processes in the body, from cancer to coronary artery disease.
Peroxisomes, on the other hand, oversee reactions that neutralize free radicals. Peroxisomes produce large amounts of the toxic H2O2 in the process, but they also contain enzymes that convert H2O2 into water and oxygen. These byproducts are safely released into the cytoplasm. Like miniature sewage treatment plants, peroxisomes neutralize harmful toxins so that they do not wreak havoc in the cells. The liver is the organ primarily responsible for detoxifying the blood before it travels throughout the body, and liver cells contain an exceptionally high number of peroxisomes.
Defense mechanisms such as detoxification within the peroxisome and certain cellular antioxidants serve to neutralize many of these molecules. Some vitamins and other substances, found primarily in fruits and vegetables, have antioxidant properties. Antioxidants work by being oxidized themselves, halting the destructive reaction cascades initiated by the free radicals. Sometimes, though, ROS accumulate beyond the capacity of such defenses.
Oxidative stress is the term used to describe damage to cellular components caused by ROS. Due to their distinctive unpaired electrons, ROS can set off chain reactions where they remove electrons from other molecules, which then become oxidized and reactive; they do the same to other molecules, causing a chain reaction. ROS can cause permanent damage to cellular lipids, proteins, carbohydrates, and nucleic acids. Damaged DNA can lead to genetic mutations and even cancer. A mutation is a change in the nucleotide sequence in a gene within a cell’s DNA, potentially altering the protein coded by that gene. Other diseases believed to be triggered or exacerbated by ROS include Alzheimer’s disease, cardiovascular diseases, diabetes, Parkinson’s disease, arthritis, Huntington’s disease, and schizophrenia, among many others. It is noteworthy that these diseases are largely age-related. Many scientists believe that oxidative stress is a major contributor to the aging process.
Aging and the Cell: The Free Radical Theory
The free radical theory on aging was originally proposed in the 1950s, and it still remains under debate. Generally speaking, the free radical theory of aging suggests that accumulated cellular damage from oxidative stress contributes to the physiological and anatomical effects of aging. There are two significantly different versions of this theory: one states that the aging process itself is a result of oxidative damage, and the other states that oxidative damage causes age-related diseases and disorders. The latter version of the theory is more widely accepted than the former. However, many lines of evidence suggest that oxidative damage does contribute to the aging process. Research has shown that reducing oxidative damage can result in a longer lifespan in certain organisms such as yeast, worms, and fruit flies. Conversely, increasing oxidative damage can shorten the lifespan of mice and worms. Interestingly, a manipulation called calorie restriction (moderately restricting the caloric intake) has been shown to increase life span in some laboratory animals. It is believed that this increase is at least in part due to a reduction of oxidative stress. However, a long-term study of primates with calorie restriction showed no increase in their lifespan. A great deal of additional research will be required to better understand the link between reactive oxygen species and aging.
The Cytoskeleton
Much like the bony skeleton structurally supports the human body, the cytoskeleton helps the cells to maintain their structural integrity. The cytoskeleton is a group of fibrous proteins that provide structural support for cells, but this is only one of the functions of the cytoskeleton. Cytoskeletal components are also critical for cell motility, cell reproduction, and transportation of substances within the cell.
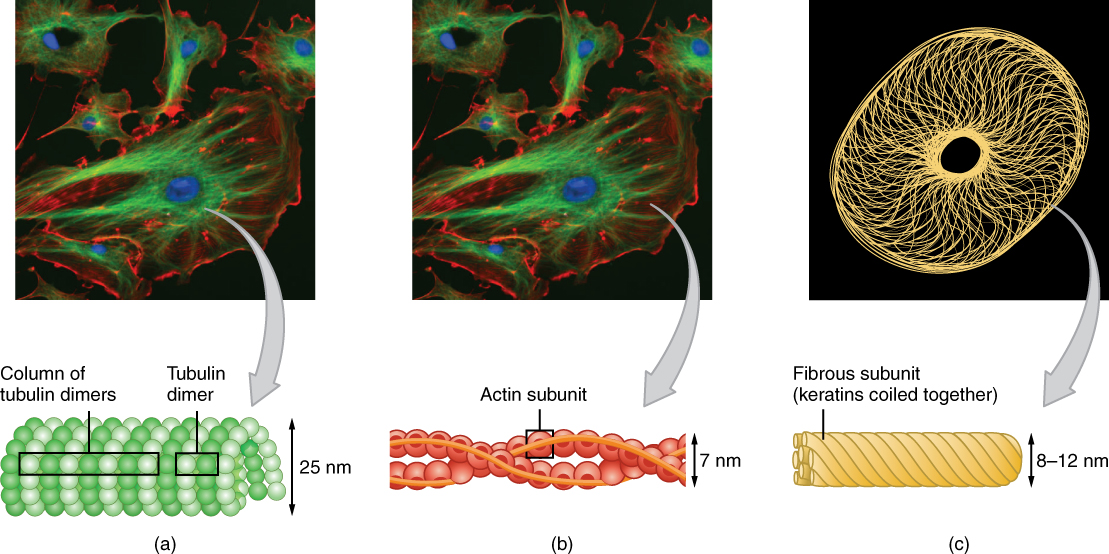
The cytoskeleton forms a complex thread-like network throughout the cell consisting of three different kinds of protein-based filaments: microfilaments, intermediate filaments, and microtubules (Figure 3.1.6). The thickest of the three is the microtubule, a structural filament composed of subunits of a protein called tubulin. Microtubules maintain cell shape and structure, help resist compression of the cell, and play a role in positioning the organelles within the cell. Microtubules also make up two types of cellular appendages important for motion: cilia and flagella. Cilia are found on many cells of the body, including the epithelial cells that line the airways of the respiratory system. Cilia move rhythmically; they beat constantly, moving waste materials such as dust, mucus, and bacteria upward through the airways, away from the lungs and toward the mouth. Beating cilia on cells in the female fallopian tubes move egg cells from the ovary towards the uterus. A flagellum (plural flagella) is an appendage larger than a cilium and specialized for cell locomotion. The only flagellated cell in humans is the sperm cell that must propel itself towards female egg cells.
A very important function of microtubules is to set the paths (somewhat like railroad tracks) along where the genetic material can be pulled (a process requiring ATP) during cell division, so that each new daughter cell receives the appropriate set of chromosomes. Two short, identical microtubule structures called centrioles are found near the nucleus of cells. A centriole can serve as the cellular origin point for microtubules extending outward as cilia or flagella or can assist with the separation of DNA during cell division. Microtubules grow out from the centrioles by adding more tubulin subunits, like adding additional links to a chain.
In contrast with microtubules, the microfilament is a thinner type of cytoskeletal filament (Figure 3.1.6b). Actin, a protein that forms chains, is the primary component of these microfilaments. Actin fibers, twisted chains of actin filaments, constitute a large component of muscle tissue and, along with the protein myosin, are responsible for muscle contraction. Like microtubules, actin filaments are long chains of single subunits (called actin subunits). In muscle cells, these long actin strands, called thin filaments, are “pulled” by thick filaments of the myosin protein to contract the cell.
Actin also has an important role during cell division. When a cell is about to split in half during cell division, actin filaments work with myosin to create a cleavage furrow that eventually splits the cell down the middle, forming two new cells from the original cell.
The final cytoskeletal filament is the intermediate filament. As its name would suggest, an intermediate filament is a filament intermediate in thickness between the microtubules and microfilaments (Figure 3.1.6c). Intermediate filaments are made up of long fibrous subunits of a protein called keratin that are wound together like the threads that compose a rope. Intermediate filaments, in concert with the microtubules, are important for maintaining cell shape and structure. Unlike the microtubules, which resist compression, intermediate filaments resist tension—the forces that pull apart cells. There are many cases in which cells are prone to tension, such as when epithelial cells of the skin are compressed, tugging them in different directions. Intermediate filaments help anchor organelles together within a cell and also link cells to other cells by forming special cell-to-cell junctions.
Section Review
The internal environment of a living cell is made up of a fluid, jelly-like substance called cytosol, which consists mainly of water, but also contains various dissolved nutrients and other molecules. The cell contains an array of cellular organelles, each one performing a unique function and helping to maintain the health and activity of the cell. The cytosol and organelles together compose the cell’s cytoplasm. Most organelles are surrounded by a lipid membrane similar to the cell membrane of the cell. The endoplasmic reticulum (ER), Golgi apparatus, and lysosomes share a functional connectivity and are collectively referred to as the endomembrane system. There are two types of ER: smooth and rough. While the smooth ER performs many functions, including lipid synthesis and ion storage, the rough ER is mainly responsible for protein synthesis using its associated ribosomes. The rough ER sends newly made proteins to the Golgi apparatus where they are modified and packaged for delivery to various locations within or outside of the cell. Some of these protein products are enzymes destined to break down unwanted material and are packaged as lysosomes for use inside the cell.
Cells also contain mitochondria and peroxisomes, which are the organelles responsible for producing the cell’s energy supply and detoxifying certain chemicals, respectively. Biochemical reactions within mitochondria transform energy-carrying molecules into the usable form of cellular energy known as ATP. Peroxisomes contain enzymes that transform harmful substances such as free radicals into oxygen and water. Cells also contain a miniaturized “skeleton” of protein filaments that extend throughout its interior. Three different kinds of filaments compose this cytoskeleton (in order of increasing thickness): microfilaments, intermediate filaments, and microtubules. Each cytoskeletal component performs unique functions as well as provides a supportive framework for the cell.
Interactive Link Questions
Watch this video to learn about the endomembrane system, which includes the rough ER, smooth ER, and the Golgi apparatus as well as lysosomes and vesicles.
Video 3.1. Nucleus and Endomembrane System by Vance Kit (Lab207)
Review Questions
Critical Thinking Questions
Glossary
- cytoplasm
- the entire contents of a cell excluding the nucleus, consisting of the cytosol and organelles
- cytosol
- the fluid portion of the cytoplasm, excluding organelles and other insoluble materials; it is a jelly-like substance within the cell that provides the fluid medium necessary for biochemical reactions
- endomembrane system
- a system of interconnected membranes within eukaryotic cells that includes the endoplasmic reticulum, Golgi apparatus, and lysosomes and is responsible for protein and lipid synthesis, modification, and transport
- endoplasmic reticulum
- a network of membranes found inside a eukaryotic cell that is involved in protein and lipid synthesis
- eukaryotic
- cells having a membrane-bound nucleus and other membrane-bound organelles
- Golgi apparatus
- an organelle found in eukaryotic cells that processes and packages proteins and lipids
- lysosomes
- organelles containing enzymes that break down and digest cellular waste and debris
- membrane
- a selective barrier; it allows sufficient passage of oxygen, nutrients, and waste to service the entire cell
- nucleus
- a membrane-bound structure in eukaryotic cells containing the cell’s genetic material (DNA) and controlling its growth and reproduction
- organelles
- specialized subunits within a cell that have specific functions, typically enclosed within their own lipid membranes
- peroxisome
- a membrane-bound organelle found in eukaryotic cells that contains enzymes involved in various metabolic reactions, including the breakdown of fatty acids and detoxification of harmful substances
- ribosome
- a complex molecular machine found within all living cells that serves as the site of protein synthesis, translating genetic code from messenger RNA into amino acid sequences
- vesicles
- small, membrane-bound sacs within a cell that store, transport, or digest cellular products and waste
Glossary Flashcards
References
Kolata, G. (2012 Aug. 29). Severe diet doesn’t prolong life, at least in monkeys. New York Times.
This work, Human Physiology, is adapted from Anatomy & Physiology by OpenStax, licensed under CC BY. This edition, with revised content and artwork, is licensed under CC BY-SA except where otherwise noted.
Images from Anatomy & Physiology by OpenStax are licensed under CC BY except where otherwise noted.
Access the original for free at OpenStax.
Image Descriptions
Figure 3.1.1. A detailed cross-sectional diagram of a eukaryotic animal cell illustrating its internal structures and organelles. The cell is roughly circular with a tan/beige cytoplasm enclosed by a brown plasma membrane. At the cell’s center is a large purple nucleus containing darker purple chromatin and a deep purple nucleolus. The nucleus is surrounded by a double-layered nuclear envelope studded with blue dots representing ribosomes, which form the rough endoplasmic reticulum extending outward in folded blue and red layers. Several orange, oval-shaped mitochondria with internal folded membranes (cristae) are scattered throughout the cytoplasm. Pink, stacked membrane layers indicate the Golgi apparatus in the lower left, with associated pink circular Golgi vesicles nearby. The cytoskeleton is represented by thin purple intermediate filaments, gray/green microtubules radiating from a small green centrosome near the cell’s right side, and reddish microfilaments distributed throughout. Additional organelles include: pink spherical lysosomes, a large pale blue vacuole at the bottom, green peroxisomes, tan smooth endoplasmic reticulum tubules, and cream-colored secretory vesicles. Small brown dots representing free ribosomes are dispersed in the cytoplasm. All structures are clearly labeled with black text and pointer lines. [Return to Figure 3.1.1]
Figure 3.1.2. A three-panel figure comparing rough and smooth endoplasmic reticulum (ER). Panel (a) is an illustrated diagram showing a purple-gray nucleus in the upper left. The rough ER extends from the nucleus as light blue flattened sacs studded with small red dots (ribosomes). The beige smooth ER appears as tubular networks without ribosomes. Light blue cylinders at upper right show cross-sections of smooth ER tubules. Panel (b) is an electron micrograph showing rough ER as parallel stacks of membrane sheets with a granular texture. Dark dots (ribosomes) line the membranes, creating the characteristic “rough” appearance. Panel (c) is an electron micrograph showing smooth ER as an interconnected network of light-colored tubules with continuous, smooth membranes lacking ribosomes. [Return to Figure 3.1.2]
Figure 3.1.3. A two-panel figure illustrating the Golgi apparatus structure and function. Panel (a) shows a diagram with the purple nucleus and blue rough ER (studded with red ribosomes) in the upper portion. Below, the peachy-pink Golgi apparatus appears as stacked, curved membrane sacs. The “cis face” (nearest the ER) receives transport vesicles budding from the rough ER, while the “trans face” (opposite side) releases secretory vesicles toward the plasma membrane. Vesicles are shown with green edges and purple contents. Panel (b) is an electron micrograph showing the Golgi as parallel stacked membranes in a curved arrangement. White circular spaces (vesicles) surround the stack. Both the “trans face” (top) and “cis face” (bottom) are labeled. Scale bar: 50 nm. [Return to Figure 3.1.3]
Figure 3.1.4. A two-panel figure illustrating mitochondrial structure. Panel (a) shows a diagram of an elongated, bean-shaped mitochondrion with a smooth beige outer membrane. The yellow inner membrane folds inward to form shelf-like cristae. The peach/orange intermembrane space separates the two membranes. Small red and green dots in the yellow matrix represent internal proteins and molecules. Panel (b) is an electron micrograph showing a mitochondrion in black and white. The dark outer membrane surrounds the organelle, while the folded inner membrane creates dark, curved cristae extending inward. The thin intermembrane space is visible between the membranes, and the lighter gray matrix has a granular texture. [Return to Figure 3.1.4]
Figure 3.1.5. A cross-sectional diagram of a virus particle shown as a sphere. At the center is a purple hexagonal structure labeled “Crystalline core” containing the viral genetic material. This core is surrounded by a yellow spherical layer. The outermost boundary is the plasma membrane, shown as a thin border with an embedded lipid bilayer. The lipid bilayer is depicted with characteristic red and gray molecular components representing the hydrophilic heads and hydrophobic tails of phospholipids arranged in a double layer. [Return to Figure 3.1.5]
Figure 3.1.6. A three-panel figure illustrating the three main types of cytoskeletal filaments, with fluorescence microscopy images above and structural diagrams below. Panel (a) shows microtubules. The top fluorescence image displays cells with blue nuclei, bright green fibrous structures (microtubules), and red cell outlines. Below, a diagram shows the microtubule structure as a hollow tube composed of green spherical tubulin dimers arranged in columns, with a 25 nm diameter. Panel (b) shows actin filaments. The fluorescence image reveals similar cellular features with red-orange threadlike actin filaments throughout the cells. The structural diagram depicts actin filaments as two intertwined strands of red-orange actin subunits forming a twisted rope structure, with a 7 nm diameter. Panel (c) shows intermediate filaments. The top image displays a circular, donut-shaped pattern of yellow filaments against a black background. The diagram illustrates intermediate filaments as rope-like cables of coiled yellow fibrous subunits (keratins wound together), with an 8-12 nm diameter. [Return to Figure 3.1.6]
Report an Error
Did you find an error, typo, broken link, or other problem in the text? Please follow this link to the error reporting form to submit an error report to the authors.
